As part of the innate immune system, interferons act to initiate the antiviral response in host organisms. While several immune cell types produce and secrete interferons (IFNs) – including natural killer cells, T cells, and macrophages – a subset of dendritic cells known as plasmacytoid dendritic cells (pDCs) are the body’s major producer of type I (α and β) interferons, though they produce type III interferons and a few key inflammatory cytokines as well (Jego et al., 2003). In general, pDCs are stimulated early in the infection cycle by the presence of viral particles, at which point they become activated and differentiate into one of three main effector types: P1, P2, or P3. Some viruses, however, have evolved to avoid or impair pDC activation, thereby preventing a successful antiviral response by the host immune system (Conry et al., 2009; Dolganiuc et al., 2006). Though the Middle East Respiratory Virus (MERS) was shown to activate pDCs in vivo, it is not currently known whether the novel coronavirus induces the same phenotypic changes. Imbalances in the cytokine and interferon profiles of COVID-19 patients has prompted investigation of the potential interaction between SARS-CoV-2 and host pDCs.
In a study from earlier this month, researchers aimed to determine the effects of SARS-CoV-2 on pDC activation and interferon secretion. First, Onodi et al. collected primary pDCs from healthy human donors; these cells were cultured for 24 hours with either SARS-CoV-2 or the influenza virus (the positive control). The authors noted significant diversification of the pDCs into the P1, P2, and P3 subsets and away from the non-activated P0 subset compared to negative controls (Figure 1). They also showed that SARS-CoV-2 interaction improved the viability of the pDCs compared to the negative control. In order to answer whether SARS-CoV-2-induced activation was dependent on infection of pDCs, they first assessed ACE2 expression on the surface of pDCs and found no significant expression compared to Vero E6 and HEK 293T-ACE2 cell lines. The pDCs were challenged with SARS-CoV-2 at a multiplicity of infection (MOI) of 2 and cultured for 2, 24, or 48 hours. The pDCs were found to be essentially refractory to infection: measurements of intracellular nucleoprotein production and release of infectious viral particles were significantly lower than in Vero E6 control cells infected with SARS-CoV-2 (Figure 1). Taken together, these data indicate that pDCs are resistant to SARS-CoV-2 infection and are activated by the virus via an ACE2-independent mechanism.
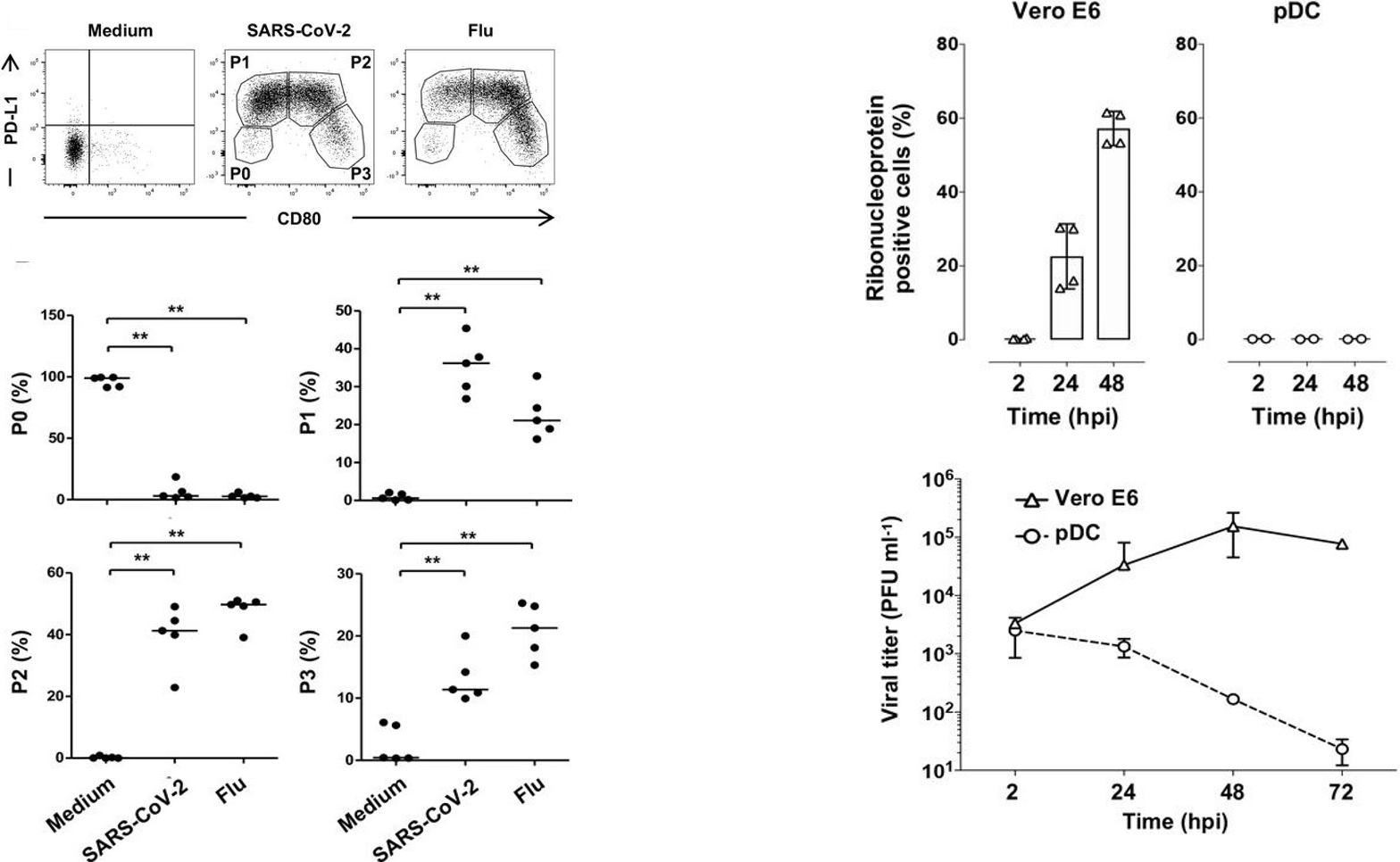
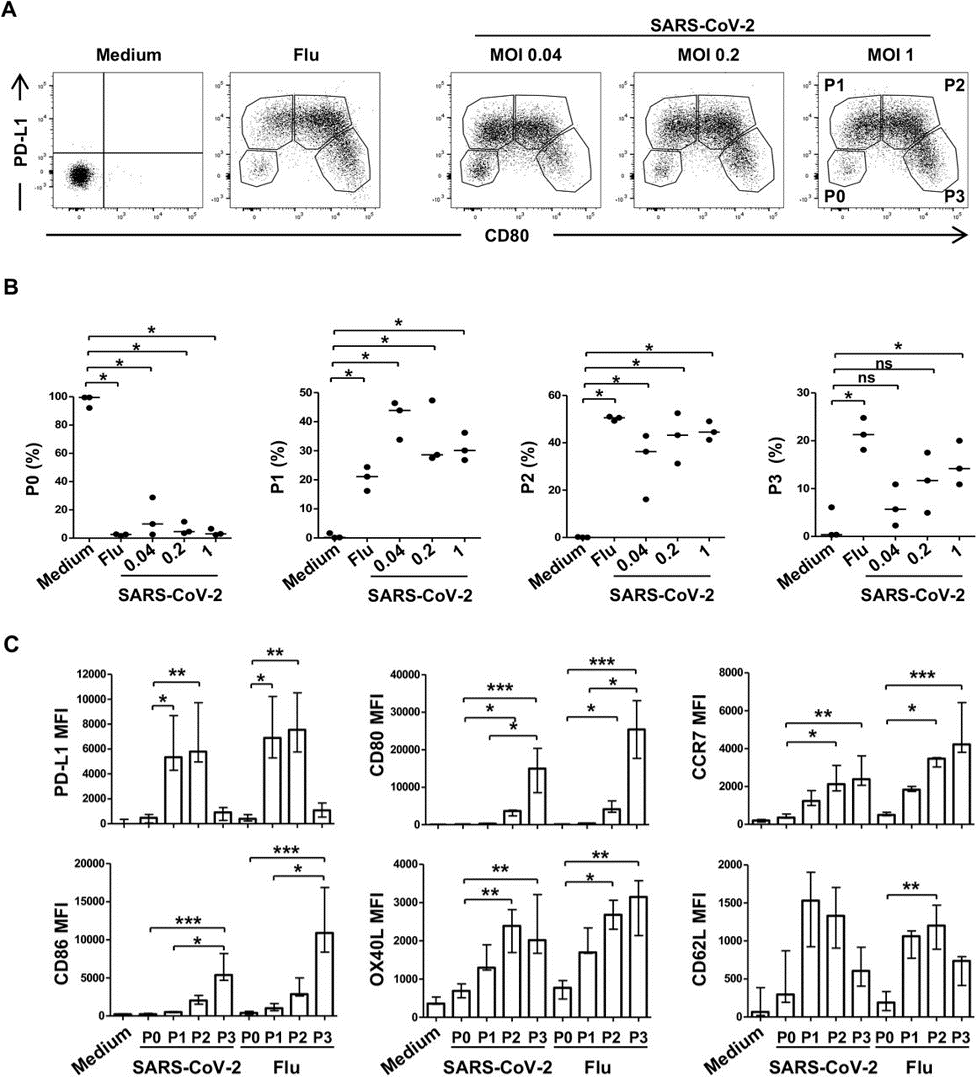
Several immune checkpoint markers are upregulated on the surface of pDCs upon activation and differentiation. These markers are critical in directing subsequent T cell stimulation and serve as markers of pDC differentiation. The authors assessed the effect of SARS-CoV-2 on checkpoint marker expression. In general, the markers CD80, CD86, CCR7, OX40L, and PD-L1 were upregulated on the cells, though specific patterns of upregulation are given in Figure 2. The upregulated expression of the checkpoint molecules persisted at significant levels at 48 hours – in particular, CD80/CD86 expression on P3-subset cells.
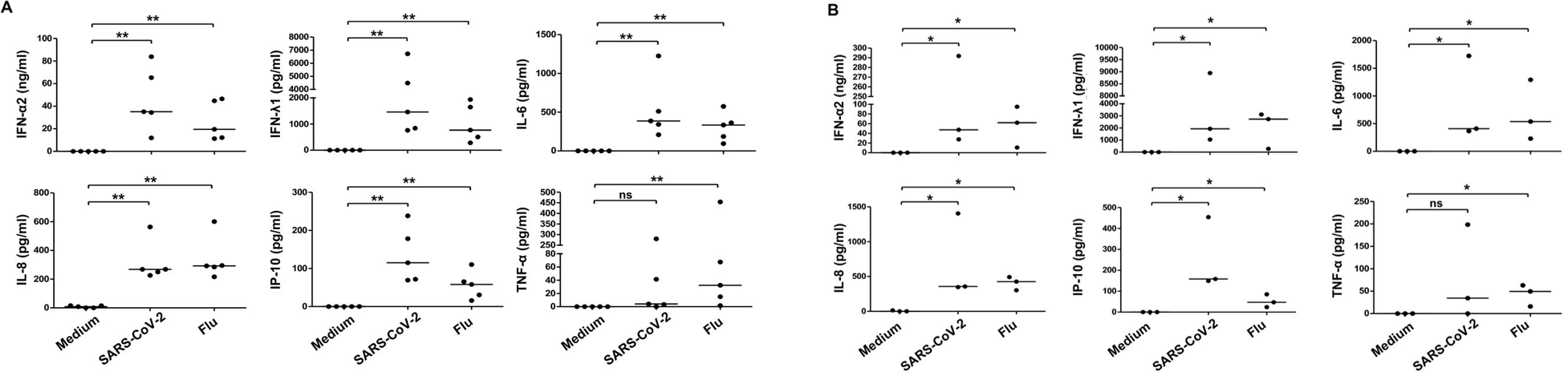
In addition to their role as the primary producers of type I interferons, pDCs also produce type III interferons, as well as a small subset of inflammatory cytokines. After 24 hours in culture with SARS-CoV-2 or the influenza virus, levels of all three – type I interferons, type III interferons, and cytokines – were significantly elevated compared to controls. Specifically, IFN-α (type I), IFN-γ (type III), the chemokine IP-10, and the cytokines IL-6 and IL-8 were all upregulated in both the SARS-CoV-2 and flu treatments. Tumor necrosis factor-α, another inflammatory cytokine, was only “marginally” induced by SARS-CoV-2. These results persisted at 48 hours in culture (Figure 3). One of the primary points of entry for SARS-CoV-2 into the human body is the oropharyngeal mucosa. Due to this fact, the authors validated their findings from peripheral pDCs using pDCs purified from tonsils. They observed significant activation and differentiation of the tonsillar pDCs into each of the three subsets; the tonsillar pDCs also produced significantly elevated IFN and cytokines, similar to peripheral pDCs.
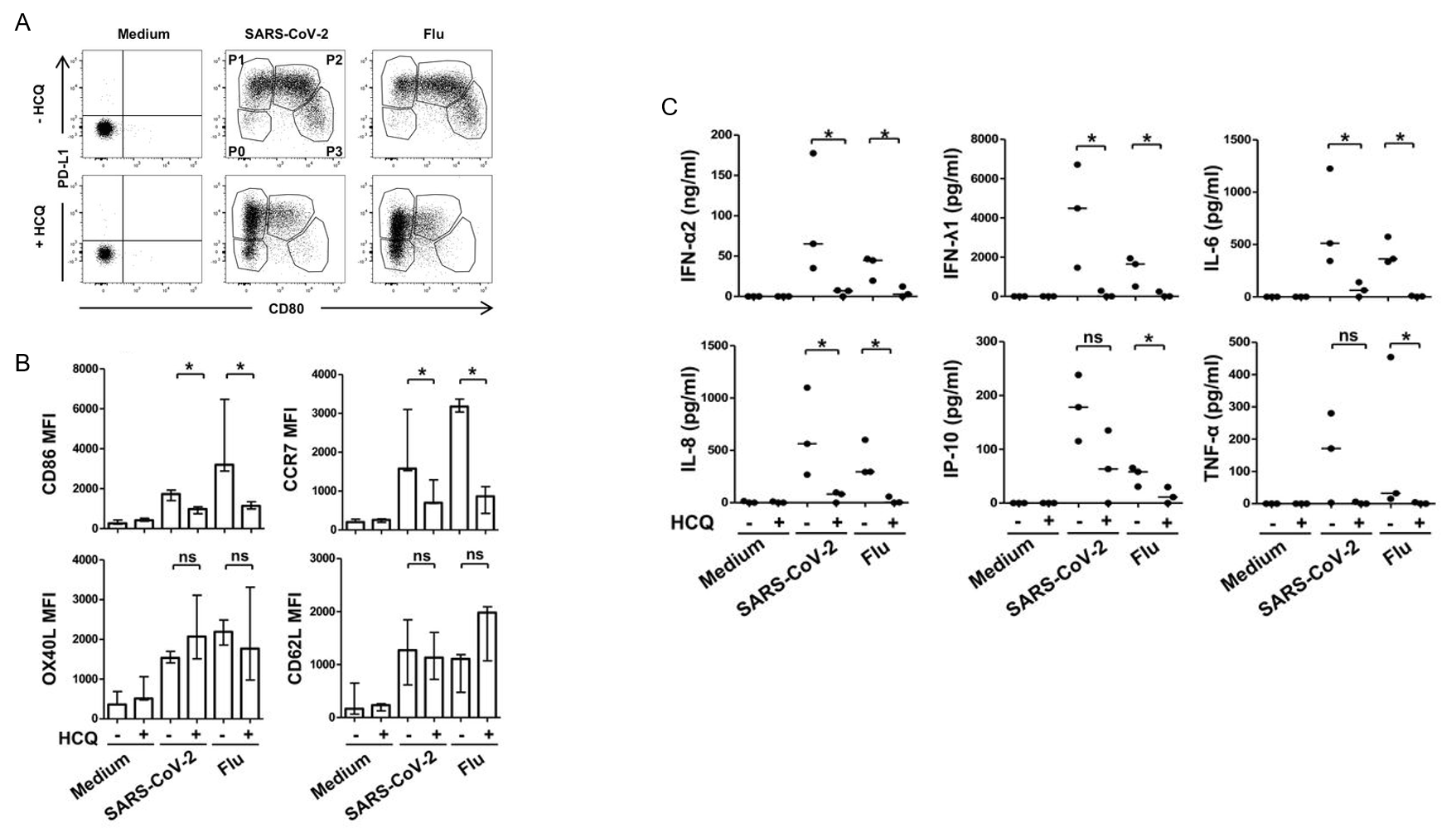
Finally, the authors hoped to determine the effects of pharmacological agents on pDC activation, differentiation, and cytokine/interferon production. Plasmacytoid DCs were cultured for 24 hours with SARS-CoV-2 in the presence or absence of hydroxychloroquine (HCQ). Post culture, pDC diversification was found to be reduced or altogether inhibited: P2 and P3 subset differentiation was almost entirely inhibited; this reduction was HCQ dose-dependent (Figure 4). The decrease in distinct subsets was accompanied by a decrease in several immune checkpoint markers, including CD80, CD86, and CCR7. There was also a significant decrease in IFN-α, IFN-γ, IL-6, and IL-8. In all, these data demonstrate that HCQ inhibits SARS-CoV-2 activation of pDCs and may actually result in adverse immune conditions for fighting against the infection.
The results outlined in this paper indicate that pDCs are not involved in or the proximal cause of the interferon/cytokine imbalance that characterizes the progression of COVID-19 to a severe state. On the contrary, interaction with SARS-CoV-2 served to enhance the production of interferons and cytokines by pDCs. The authors note that these outcomes suggest that the immunological defects present in severe and critical COVID-19 patients are “acquired during disease evolution through secondary events”, which may not be directly associated with the virus itself. This work paves the way for future investigation of the explicit interaction between SARS-CoV-2 and pDCs, and of the cause of cytokine and interferon imbalances. Additionally, this work highlights the detrimental effects of hydroxychloroquine. Though this work has not yet been peer reviewed, it is one paper in a growing catalogue espousing caution in regard to the use of this drug for treatment of SARS-CoV-2 infection.
References
Conry, S. J., Milkovich, K. A., Yonkers, N. L., Rodriguez, B., Bernstein, H. B., Asaad, R., Heinzel, F. P., Tary-Lehmann, M., Lederman, M. M., & Anthony, D. D. (2009). Impaired Plasmacytoid Dendritic Cell (PDC)-NK Cell Activity in Viremic Human Immunodeficiency Virus Infection Attributable to Impairments in both PDC and NK Cell Function. Journal of Virology, 83(21), 11175–11187. https://doi.org/10.1128/jvi.00753-09
Dolganiuc, A., Chang, S., Kodys, K., Mandrekar, P., Bakis, G., Cormier, M., & Szabo, G. (2006). Hepatitis C Virus (HCV) Core Protein-Induced, Monocyte-Mediated Mechanisms of Reduced IFN-α and Plasmacytoid Dendritic Cell Loss in Chronic HCV Infection. The Journal of Immunology, 177(10), 6758–6768. https://doi.org/10.4049/jimmunol.177.10.6758
Jego, G., Palucka, A. K., Blanck, J. P., Chalouni, C., Pascual, V., & Banchereau, J. (2003). Plasmacytoid dendritic cells induce plasma cell differentiation through type I interferon and interleukin 6. Immunity, 19(2), 225–234. https://doi.org/10.1016/S1074-7613(03)00208-5
Onodi, F., Bonnet-Madin, L., Karpf, L., Meertens, L., Poirot, J., Legoff, J., Delaugerre, C., Amara, A., & Soumelis, V. (2020). SARS-CoV-2 induces activation and diversification of human plasmacytoid pre-dendritic cells. BioRxiv, 2020.07.10.197343. https://doi.org/10.1101/2020.07.10.197343
Leave a Reply