Thus far in the pandemic, we’ve seen an array of clinical presentations of COVID-19, including reports of SARS-CoV-2-specific T cell responses, severe lymphopenia, cytokine storm, not to mention the swath of completely asymptomatic or mildly symptomatic individuals (Huang et al., 2020; Ju et al., 2020; Kuri-Cervantes et al., 2020; Laing et al., 2020, Yang, Gui, Xiong, 2020). Clearly, better understanding of the disease course and its varied manifestations is necessary for more efficient treatment of COVID-19 patients. A more substantial grasp of the phasic progression of the disease may also enable more proactive mitigation of symptoms and allow prediction of disease progress and ultimate severity. In this article, Laing et al. advocate for the identification of common immune phenotypes – including cytokines, lymphocyte and leukocyte subsets, antibody production, etc. – in COVID-19 patients that comprise a “core disease-associated signature”. Identification of such a signature would allow for strong and clinically meaningful predictions of disease severity among these patients. They note, however, the challenges in determining any sort of consensus immune signature presented by the extensive heterogeneity of the patient population in regard to sex, age, ethnicity, and underlying comorbidities, among others. Despite this, the authors set out to characterize the immune signature of COVID-19.
63 patients with PCR-positive COVID-19 were recruited from two English hospitals; blood samples were collected from each at three distinct timepoints: first within 24 hours of recruitment, then at day 3 and then day 9 post recruitment. In addition to these patients, 10 patients with unspecified, non-COVID-19 lower respiratory tract infections were recruited as controls. These patients repeatedly tested negative for SARS-CoV-2 over the course of the study. Finally, 55 healthy volunteers were recruited as controls. These healthy controls were broken into seropositive and seronegative controls – seropositive controls having previously experienced either asymptomatic or mild infection, in neither case requiring hospitalization.
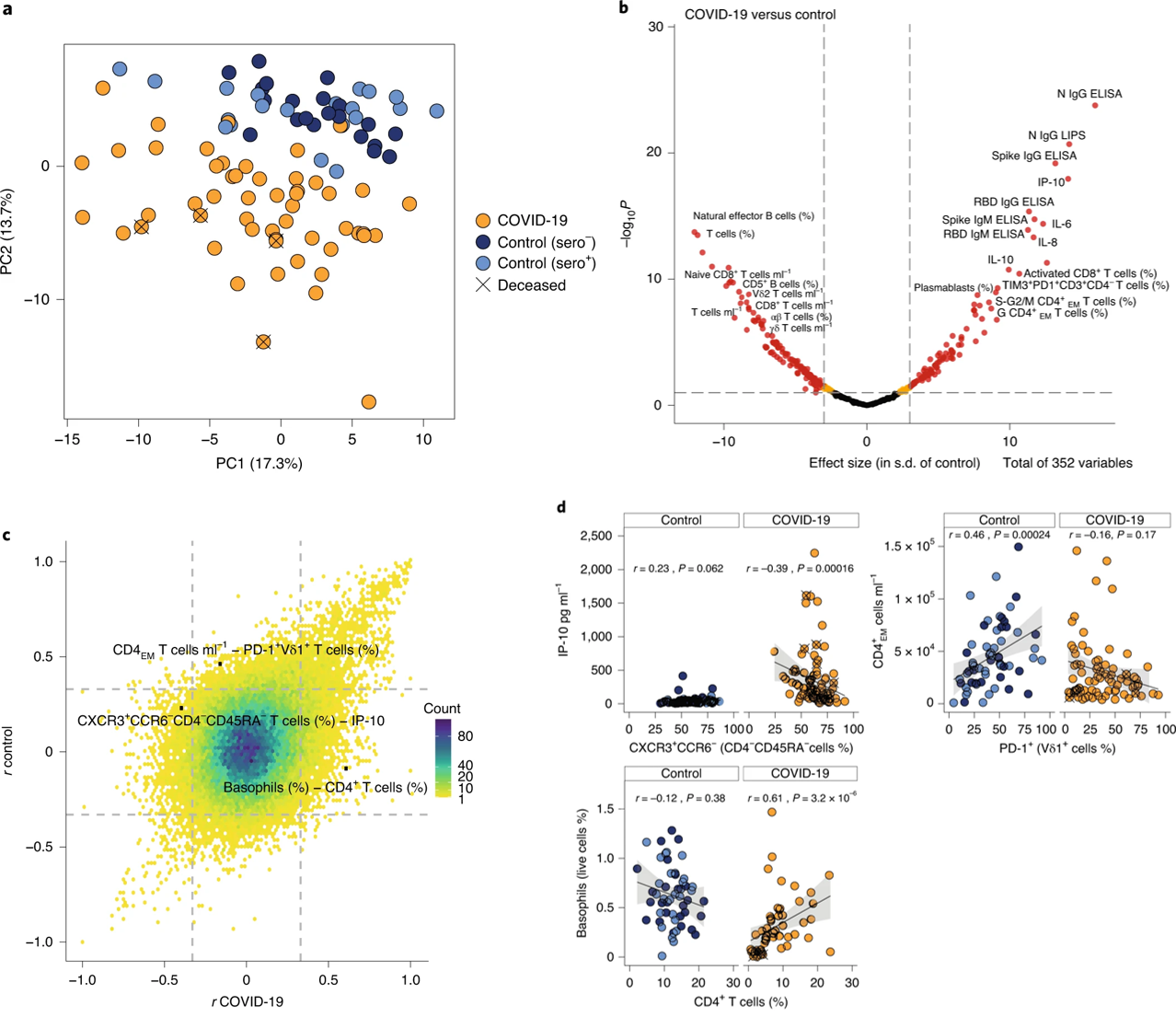
The authors began with a high-level overview of the immunological phenotypes of all subjects. They utilized eight broad multiparameter flow cytometry panels to observe a total of 176 specific phenotypes, where a single phenotype may be the count of anti-N protein IgG or CD5+ B cells, for example. A principal component analysis of these 176 phenotypes from each patient based solely on the flow cytometry parameters they established segregated COVID-19 patients from both seropositive and seronegative healthy controls (Figure 1a). To determine which of the phenotypes tested most strongly discriminated COVID-19 patients from controls, the authors next analyzed the effect size for each one (Figure 1b). They noted that factors such as SARS-CoV-2-specific antibodies, plasmablasts, CD8+ T cells expressing the exhaustion markers PD-1 and TIM3, and the cytokines IL-8, IL-10, and IL-6, and the chemokine IP-10 were prominent in the immunophenotype of COVID-19 patients compared to controls. Next, they calculated correlations between each phenotype for COVID-19 patients and plotted the coefficient for each against the coefficient for the equivalent correlation in the healthy controls (Figure 1c). Most of these correlations did not significantly differ between the populations. A subset of these correlations are given in Figure 1d. The authors suggest that these correlations compose an “aggregate immune structure” of COVID-19 that may be used to distinguish it from other similar pathologies.
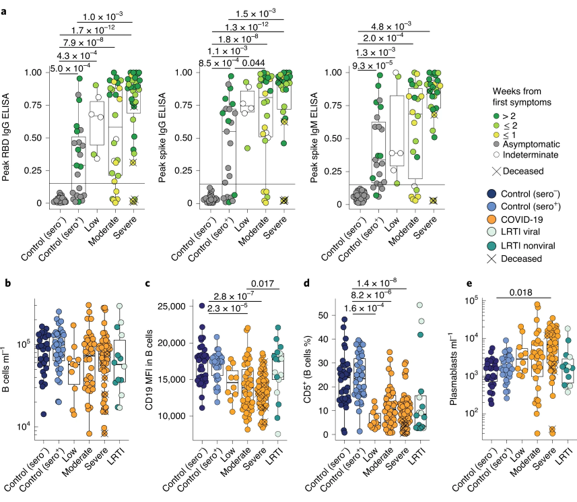
Once they had established a high-level immunophenotype of COVID-19, the authors then began analyzing several aspects of this signature in detail, seeking phenotypes that correlated with severity. They began with antibodies and the consensus B cell response. Analysis of SARS-CoV-2 RBD-specific IgG and SARS-CoV-2 Spike protein-specific IgG and IgM revealed a clear trend toward higher antibody titers in the severe cases, though this trend was not significant (Figure 2a). They note that this trend may simply reflect the fact that, in general, severe patients have had more time than controls or low-severity patients to develop an antibody response. Observation of the whole B cell repertoire showed little correlation with severity, or even with COVID-19 relative to healthy controls (Figure 2b). However, they did record significant downregulation of CD19 and depletion of CD5+ B cells in COVID-19 patients, both of which correlated with severity (Figure 2c, d). Unsurprisingly, COVID-19 patients showed increased CD38+CD27+ plasmablast frequencies, which is in line with the observed increase in antibody titers associated with disease progression (Figure 2e). Of the phenotypes observed, only plasmablast frequency was significantly correlated with severity of disease.
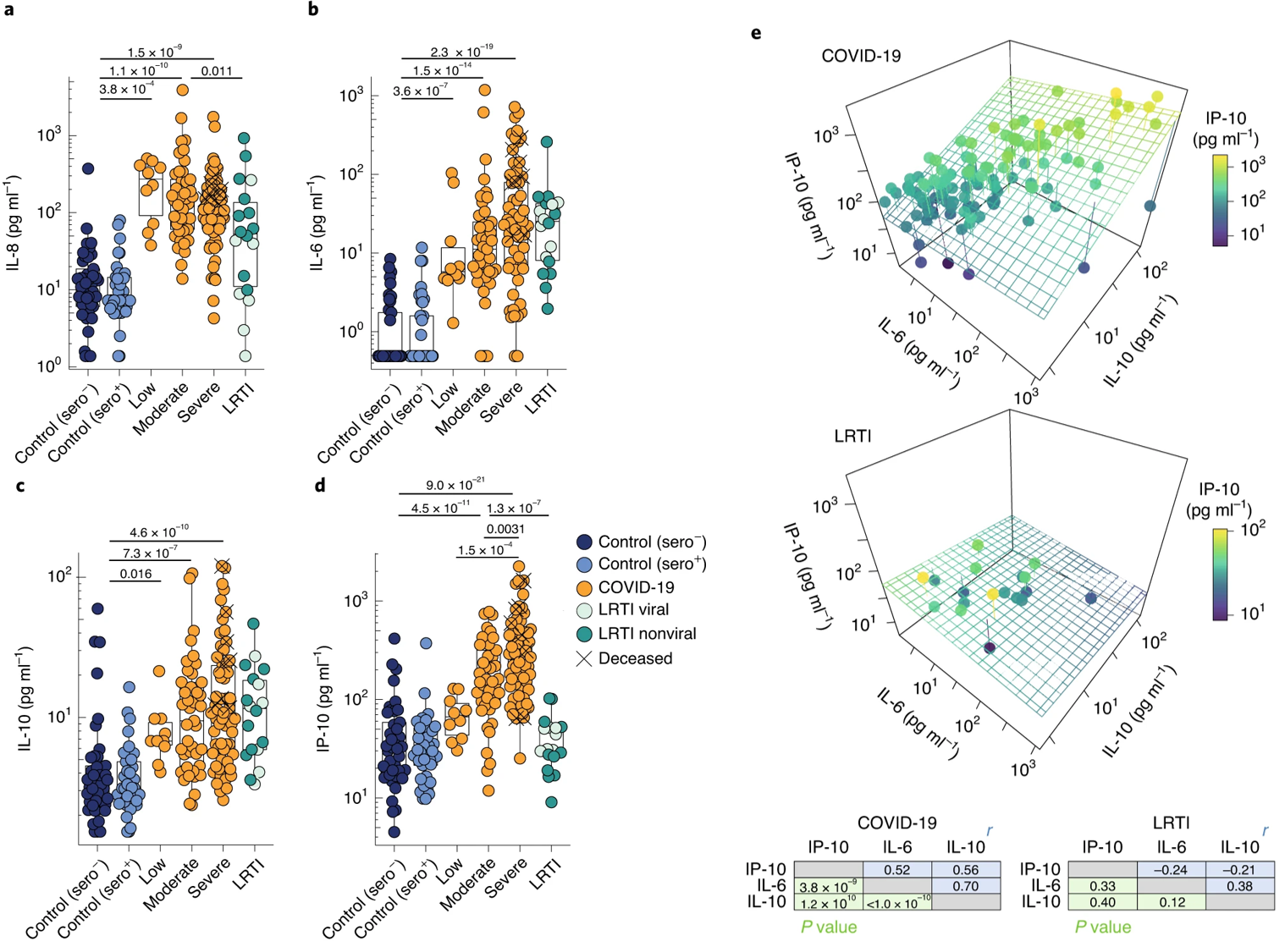
Next, the authors investigated the profiles of cytokines commonly associated with infections. IL-8, IL-6, IL-10, and the chemokine IP-10 were all significantly elevated in COVID-19 (and most were elevated in LRTI patients) compared to healthy controls (Figure 3a-d). However, only IL-6, IL-10, and IP-10 were significantly correlated with disease severity (Figure 3b-d). The authors deem this the “severity-related triad” and return to it throughout as a significant marker of disease severity and predictor of disease course. Indeed, many patients with COVID-19, but not LRTIs, were described by this triad (Figure 3e).
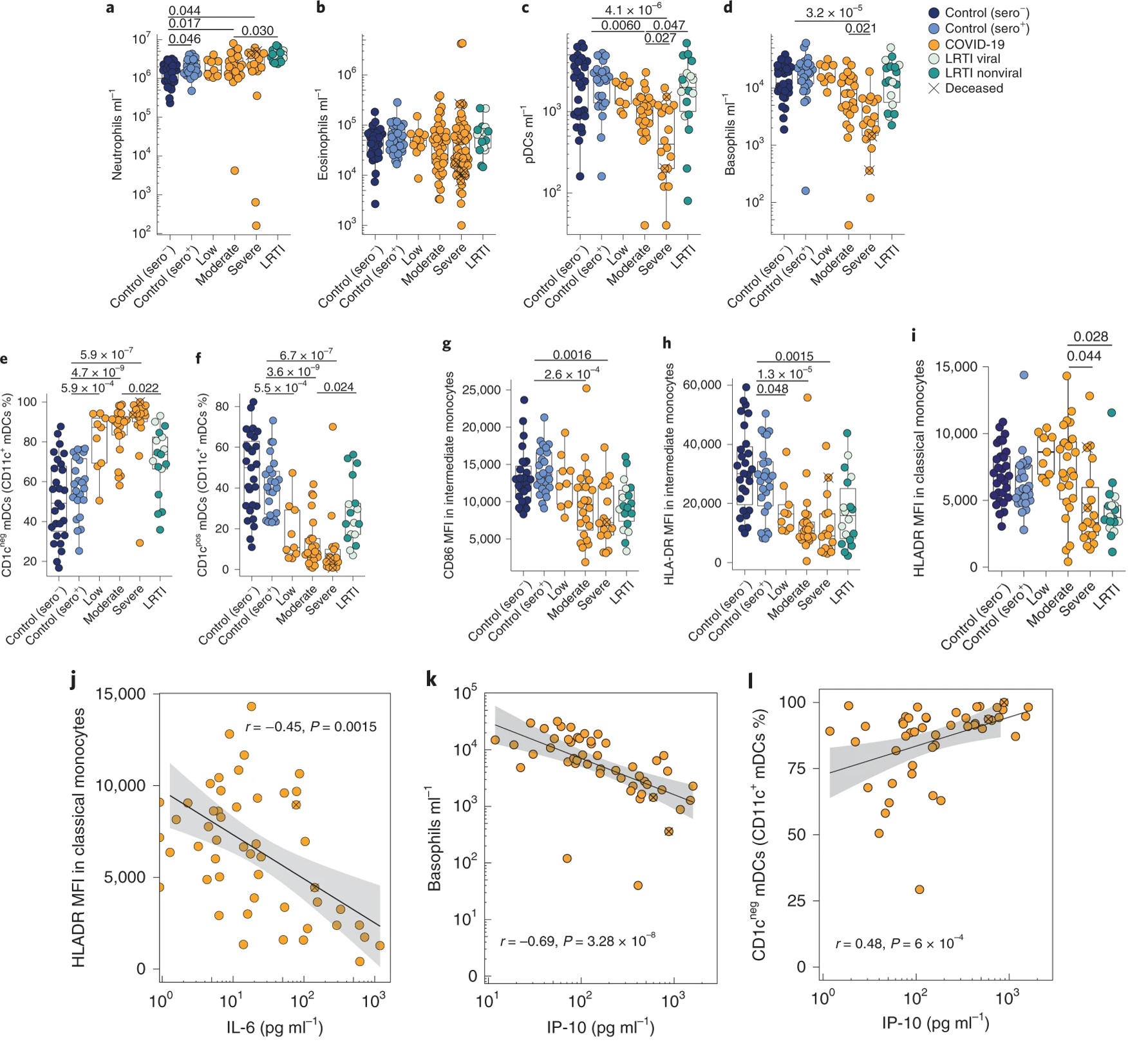
Following their analysis of cytokines and chemokines, the authors hypothesized that overexpression of certain cytokines may be reflected in the immune cell composition of these patients. IL-8, for example, activates neutrophils in vivo, and their counts were slightly elevated in COVID-19 patients, though only moderately so (Figure 4a). Eosinophils, on the other hand, displayed no significant upregulation in COVID-19 or LRTI compared to healthy controls (Figure 4b). Plasmacytoid dendritic cells and basophils were significantly and dramatically depleted in COVID-19, and their depletions correlated with severity (Figure 4c, d). Neither, however, was depleted in non-COVID-19 LRTIs. In the blood, CD11c+CD1c- dendritic cells became the predominant subset, with concomitant depletions of the CD11c+CD1c+ subset (Figure 4e, f). Of the monocyte population, the most overt phenotype was an almost universal depletion of CD86 and HLA-DR expression, especially in intermediate monocytes, though this was also observed in classical monocyte populations (Figure 4g-i). This depletion was not observed in LRTIs (Figure 4g-i). The authors then drew correlations between immune cell phenotypes and patterns of cytokine expression observed in COVID-19. IL-6 upregulation was correlated with HLA-DR downregulation (Figure 4j), IP-10 upregulation was similarly correlated with basophil downregulation and with CD1c- dendritic cell upregulation (4k, l).
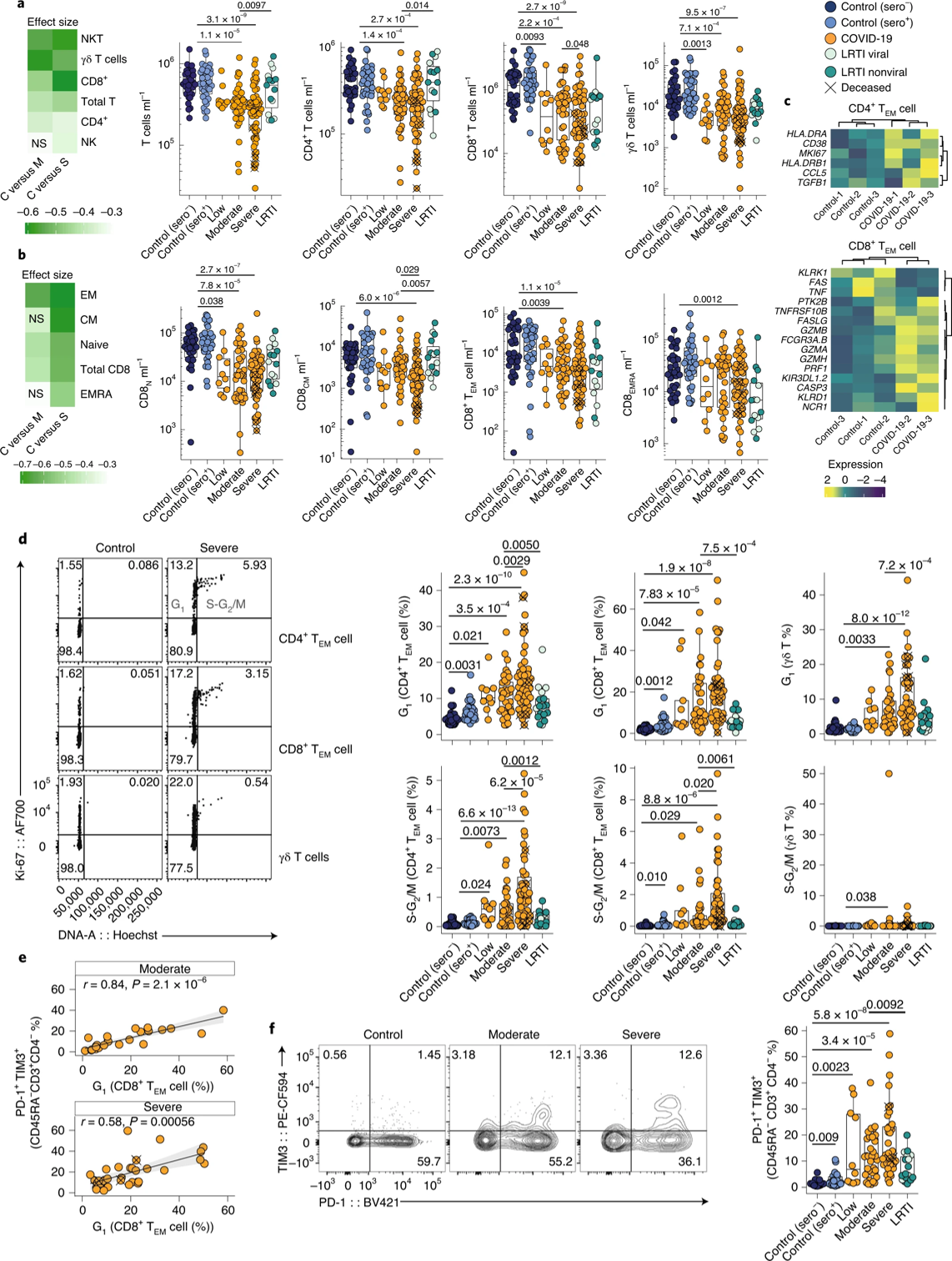
Given that some T cell phenotypes had the largest effect size in discriminating the COVID-19 population from healthy controls, they next sought to characterize the T cell signature of COVID-19. The most significant and overt effect on the T cell population was a depletion of CD8+ and γδ T cells, though depletions were also present in CD4+ T cells and in the overall T cell population (Figure 5a). Indeed, CD8+ and γδ T cells had the largest effect size – of the T cell subsets evaluated – in discriminating COVID-19 patients from healthy controls (Figure 5a). The authors also analyzed the effector memory subsets of T cells and found that CD8+ TEM were depleted in COVID-19 and correlated with severity (Figure 5b). Naïve (N) and central memory (CM) T cells were particularly downregulated in these patients, especially in severe cases (Figure 5b). Gene expression analysis in CD4+ and CD8+ T cells revealed marked upregulation of HLADR, CD38, and MK167 in CD4+ T cells, and of NCR1, FASLG, and PRF1 in CD8+ T cells, suggesting heightened activation and cell cycling among these cell subsets (Figure 5c). Additionally, the authors noted a roughly tenfold increase in the number of γδ T cells, and CD4+ and CD8+ TEM cells in the G1 or S-G2/M phase of the cell cycle (Figure 5d). This is surprising, given the fact that normally only 3% of T cells are present in these phases. The frequency of CD8+ T cells in the G1 phase also correlated with the expression of the exhaustion markers PD-1 and TIM3, and this upregulation was correlated significantly with severity of the disease (Figure 5e, f).
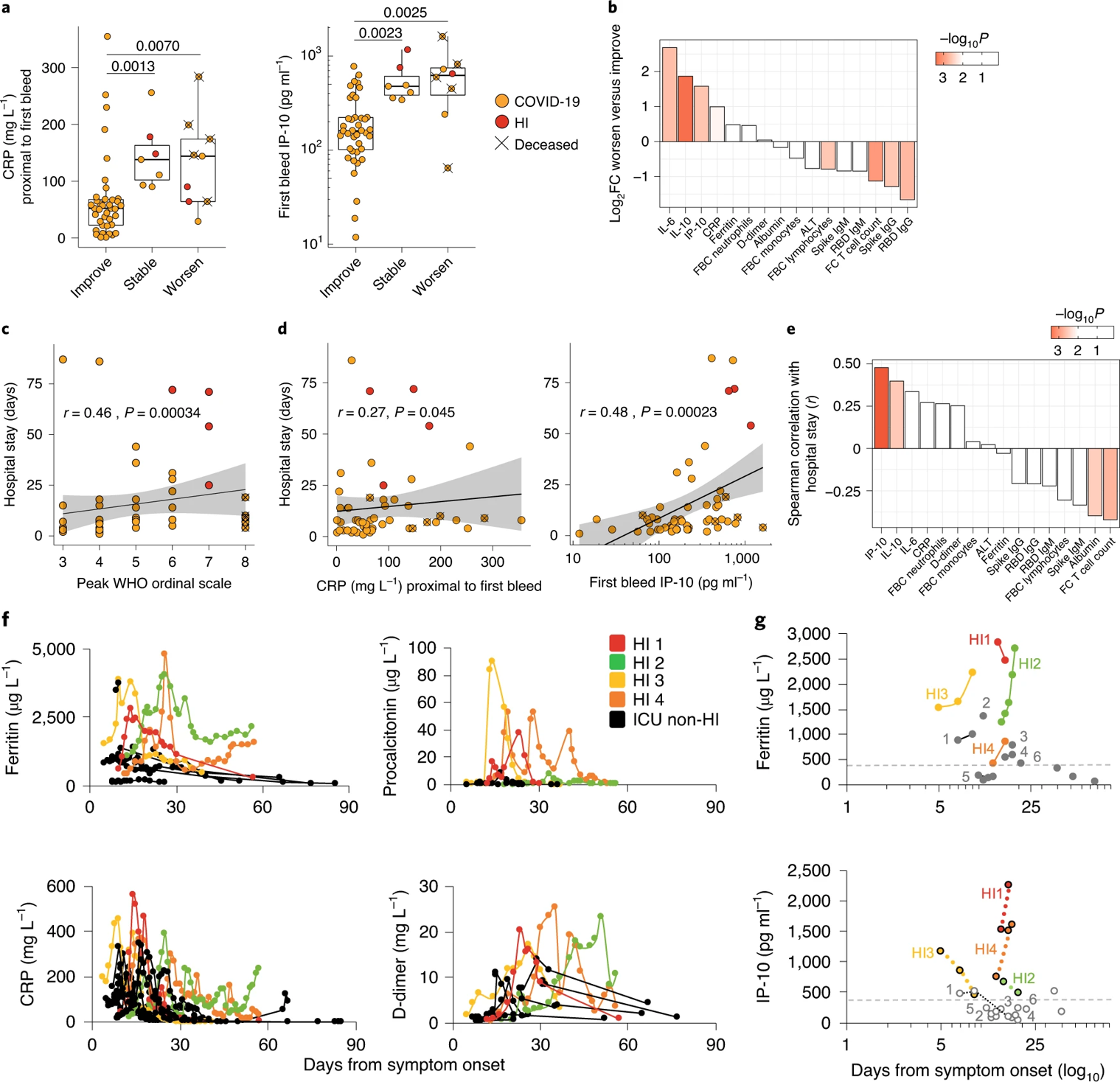
The authors express interest in, and the importance of, stratification of patients early on into those more likely to recover rapidly – this in an attempt to make the distribution and allocation of hospital resources more efficient. To this end, the authors compared the first-bleed traits in patients segregated according to whether their WHO ordinal scores decreased, increased, or stabilized during the week following admission. They note that their immunological triad of IL-10, IL-6, and IP-10 showed a greater capacity than C-reactive protein (CRP) to segregate patients based on who would improve and who would worsen (Figure 6a, b). They emphasize that measurements like anti-SARS-CoV-2 IgG and flow-cytometric T cell quantification better anticipated improvement than the canonical markers of disease severity and progression (clinical lymphocyte count, neutrophil count, ferritin, D-dimer, albumin). They found that the length of a patient’s hospital stay correlated to a small degree with peak ordinal severity, but not well with CRP (Figure 6c, d). However, length of stay was best estimated by IP-10 and IL-10 and reciprocal of T cell counts (Figure 6d, e). Finally, the authors note the ability of IP-10 to segregate hyperinflammatory patients (those with persistent fevers and hypoxemia, hyper-ferritinemia, and high CRP) from other patients requiring ICU care, above and beyond the abilities of ferritin, procalcitonin, CRP, and D-dimer (Figure 6f, g).
This paper performed the very important task of conglomerating many of the data points people have been observing and recording into a single analysis, such that they could be compared against one another. This work highlights the key role and viability of a consensus immune signature in COVID-19, which can be used to drive medical decision making. In all, the analysis performed here may potentially set the stage for new methodologies in treatment and the allocation of hospital resources, as least in terms of COVID-19 patients.
References
Huang, C., Wang, Y., Li, X., Ren, L., Zhao, J., Hu, Y., Zhang, L., Fan, G., Xu, J., Gu, X., Cheng, Z., Yu, T., Xia, J., Wei, Y., Wu, W., Xie, X., Yin, W., Li, H., Liu, M., … Cao, B. (2020). Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. The Lancet, 395(10223), 497–506. https://doi.org/10.1016/S0140-6736(20)30183-5
Ju, B., Zhang, Q., Ge, J., Wang, R., Sun, J., Ge, X., Yu, J., Shan, S., Zhou, B., Song, S., Tang, X., Yu, J., Lan, J., Yuan, J., Wang, H., Zhao, J., Zhang, S., Wang, Y., Shi, X., … Zhang, L. (2020). Human neutralizing antibodies elicited by SARS-CoV-2 infection. Nature, 584(7819), 115–119. https://doi.org/10.1038/s41586-020-2380-z
Kuri-Cervantes, L., Pampena, M. B., Meng, W., Rosenfeld, A. M., Ittner, C. A. G., Weisman, A. R., Agyekum, R. S., Mathew, D., Baxter, A. E., Vella, L. A., Kuthuru, O., Apostolidis, S. A., Bershaw, L., Dougherty, J., Greenplate, A. R., Pattekar, A., Kim, J., Han, N., Gouma, S., … Betts, M. R. (2020). Comprehensive mapping of immune perturbations associated with severe COVID-19. Science Immunology, 5(49). https://doi.org/10.1126/sciimmunol.abd7114
Laing, A. G., Lorenc, A., del Molino del Barrio, I., Das, A., Fish, M., Monin, L., Muñoz-Ruiz, M., McKenzie, D. R., Hayday, T. S., Francos-Quijorna, I., Kamdar, S., Joseph, M., Davies, D., Davis, R., Jennings, A., Zlatareva, I., Vantourout, P., Wu, Y., Sofra, V., … Hayday, A. C. (2020). A dynamic COVID-19 immune signature includes associations with poor prognosis. Nature Medicine, 1–13. https://doi.org/10.1038/s41591-020-1038-6
Yang, R., Gui, X., & Xiong, Y. (2020). Comparison of Clinical Characteristics of Patients with Asymptomatic vs Symptomatic Coronavirus Disease 2019 in Wuhan, China. JAMA Network Open, 3(5), e2010182. https://doi.org/10.1001/jamanetworkopen.2020.10182
Leave a Reply